
8-900-374-94-44

Стабильно работающая сигналка автомобильная Аллигатор подходит для установки на машины разных годов выпуска. Для питания необходимо напряжение 12 В и величина тока не больше 20 А. Контроль параметров работы осуществляют благодаря пульту-пейджеру, который комплектуется 1,5 В батарейкой.
В моделях охранной сигнализации, выпущенных после 2021 года, установлены дополнительные блоки управления, коммутаторы. Они обеспечивают взаимодействие с иными защитными модулями, но при этом требуют установки еще одного источника питания, который потребляет 3 В. Водитель не спешит с установкой, потому что вначале обсуждают детали с представителем автомобильного салона. Он расскажет, какой набор опций требуется отдельно взятой машине.
На заметку!
Не монтируют охранную систему подобного класса на грузовики и на мотоциклы. Причина заключается в том, что здесь схема подключения охранной сигнализации Аллигатор не выдержит напряжения в 24 В.
Акулы капиталистического бизнеса очень остро чувствуют зарождение нового рынка сбыта. В середине 90-х годов прошедшего века, во времена развитого бандитского капитализма в стране появилась американская Выбор её интересов распространялся на автомобильную и бытовую электронику. В том числе и на выпуск лучших автомобильных сигнализаций – дебют за новинкой 1997 года Alligator L300.
Так зарождалось российское представительство этого производителя — закрытое акционерное общество «Сатурн Хай-Тек» (с 2013 г. АО «). Оно то и стало продвигать на рынках СНГ все лучшие новинки американцев. В их обзоре был и высоколиквидный товар защиты от угонов, под настораживающим никнеймом «Alligator».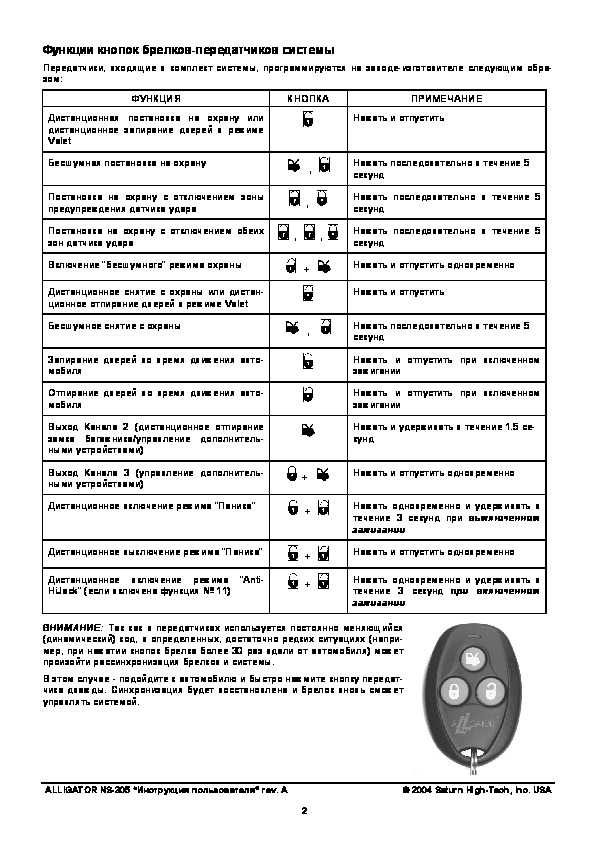 Официальный сайт бренда автосигнализации Alligator https://alligator-alarms.ru.
Официальный сайт бренда автосигнализации Alligator https://alligator-alarms.ru.
Количество охранных зон, каждая из которых функционирует независимости или связано. Рекомендована установка автомобильной сигнализации Аллигатор с 4-6 независимыми контрольными точками. Если злоумышленник выключить 1-2 охранных зоны, это не снизит уровень защиты машины. Функционал брелока – бюджетные модели снабжены кнопкой включения и выключения. В защитных системах, которые относят к среднему и к высокому ценовому диапазону, брелок снабжен LED экраном, отображающим параметры работы. Перечень других опций:
 Показатель колеблется от 1 до 2,5 км.
Показатель колеблется от 1 до 2,5 км.Функциональная сигнализация аллигатор снабжена брелоком, который не допускает случайного нажатия кнопок. В описании производитель указывает, что опция блокировка входит в комплект. Защиту от радиоэлектронного излучения, применяемого для взлома машины, обеспечивает цифровое или встроенное реле.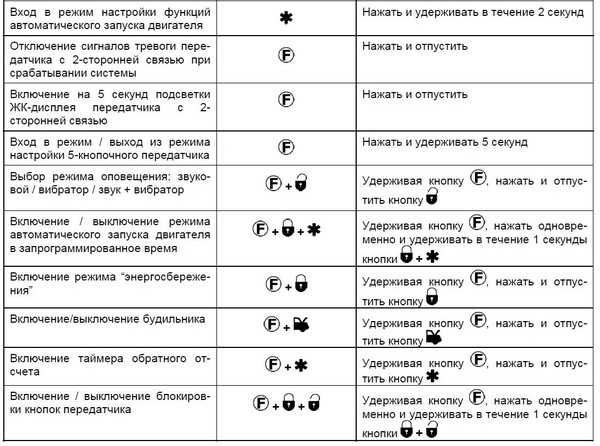 Информация о его наличии доступна в инструкции по эксплуатации. Фирменная сигнализация для машин Alligator включает ряд других возможностей:
Информация о его наличии доступна в инструкции по эксплуатации. Фирменная сигнализация для машин Alligator включает ряд других возможностей:
Нет предела для дальнейшего совершенства. Разработанная автосигнализация среднего диапазона Alligator оснащена дополнительными выходами, которые нужны для подключения новых защитных блоков.
На заметку! В инструкции производитель указывает, что система оснащена опцией подключения не менее 2-х дополнительных каналов связи. Если подобной отметки нет, то новые защитные блоки не получится установить.
Установленная сигнализация Аллигатор инструкция к которой содержит исчерпывающие сведения, представлена в 2-х модификациях. Бюджетный вариант без автоматического запуска и модель с таковым. Упомянутая опция полезна зимой, когда требуется заблаговременно прогреть двигатель. Не обойтись без автозапуска лет. Водитель, нажав соответствующую кнопку на брелоке, активирует климат-контроль. К моменту его прихода в салоне будет комфортная температура. Установленная сигнализация Аллигатор с автозапуском может быть активирована по одному из критериев, перечисленных ниже:
Система Аллигатор с установленным автозапуском представлена в нескольких модификациях. Вначале сравнивают характеристики каждой из них, а потом заказывают установку.
Вначале сравнивают характеристики каждой из них, а потом заказывают установку.
| Модель | Доступные опции | Радиус действия (км) | Примечание |
| C 500 | Обратная связь отображает параметры работы двигателя и состояние защиты, иммобилайзер, защита от перехвата радиосигнала | 2,5 на открытой местности | Удаленный запуск только по таймеру |
| S 325 | Пульт с LED экраном обеспечивают контроль и настройку параметров работы, | 1,6-1,8 | Установка возможна на машины всех возрастов |
| S 875 RS | Обратная связь, блокировка клавиш брелока и силового агрегата, наличие возможное для расширения | 2 на открытой местности | Действующая схема автомобильной сигнализации Аллигатор включает 4 датчика удара |
| S 250 | 3 кнопки управления с блокировкой, световая индикация режимов | 1,8 | Наличие защиты от ложных включений, доступна система вибрации брелока управления |
| L 530 | Антихайджек, дополнительный канал передачи радиосигнала | 1,8 | Поставляемый в комплекте брелок Аллигатор 4 удобных кнопки оснащен возможностью управления оптикой, стеклоподъемниками, дверью багажника |
| PS 302 | Брелок с опцией блокировки кнопок | 0,6 в городской среде; 1,8 на открытой местности | – |
Бюджетная сигнализация Аллигатор инструкция по применению и брелок к которой идут в комплекте, оснащена лишь кнопками включения, открытия багажника.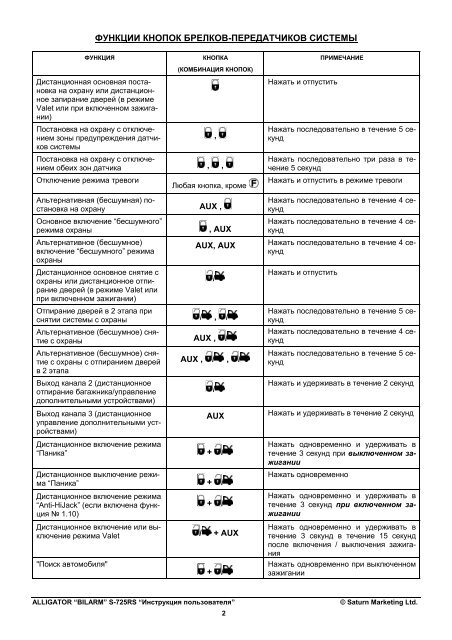 Отсутствие опции автозапуск компенсируется более низкой стоимостью обслуживания:
Отсутствие опции автозапуск компенсируется более низкой стоимостью обслуживания:
A light – односторонняя система связи с возможностью блокировки двигателя. В отличие от двух перечисленных выше разновидностей, A light имеет защиту от устройств-сканеров.
На заметку! Держа в руках брелок управления, получится сразу понять, можно или нет включить автозапуск на автомобильной сигнализации Аллигатор. Наличие литеры «М» говорит об отсутствии не только упомянутой опции, но и 2-х сторонней связи. Класс «М», в отличие от «А» и «C2», обладает меньшим функционалом.
Светодиодный индикатор обычно располагают в одном из верхних углов лобового стекла. Индикатор обычно крепится при помощи двухстороннего скотча. Ноесли хочется закрепитьэлемент на приборной панели, то понадобится предварительно сверлить в ней отверстие диаметром в 7мм.
Ноесли хочется закрепитьэлемент на приборной панели, то понадобится предварительно сверлить в ней отверстие диаметром в 7мм.
Датчик удара лучше всего расположить на твердой поверхностипереборки, которая отделяет салон от двигательного отсека.Установка этого элемента сигнализации Аллигатор осуществляется при помощи 2-х винтов, помимо этого, прибор необходимо так разместить, чтобы потом можно было добраться до его регулировочных резисторов. Весьма важно правильно расположить датчик температуры Аlligator.
Лучше всего датчик температуры закреплять под капотом. Обязательно стоит учитывать, что датчик температуры должен крепиться к резиновой части радиатора.
Это обусловлено тем, что этот сенсор может реагировать не только на изменение температуры охлаждающей жидкости, но и на показатели металлических элементов. Крепиться датчик температуры при помощи стяжек.
Модуль приёма-передачи Аlligatorжелательно установить в области лобового стекла. Это необходимо для того чтобы дальность связи была максимальной. Ни в коем случае не стоит устанавливать этот модуль на боковые стойки, так как воздействие сильной вибрации может нарушить работу данного устройства. В случае если лобовое стекло обработано отражающим покрытием, датчик Аlligatorможно разместить в задней части авто. Приемопередающий датчик крепится при помощи липкой ленты.
Ни в коем случае не стоит устанавливать этот модуль на боковые стойки, так как воздействие сильной вибрации может нарушить работу данного устройства. В случае если лобовое стекло обработано отражающим покрытием, датчик Аlligatorможно разместить в задней части авто. Приемопередающий датчик крепится при помощи липкой ленты.
Оригинальная сигнализация автомобильная Аллигатор инструкция по эксплуатации к которой входит в комплект поставки, должна быть подобрана с учетом возраста машины, его электрической цепи и модели. Учитывают советы завода изготовителя. Быстрая настройка машинной сигнализации Аллигатор возможна в авторизован центре СТО. Здесь расскажут о «+» и о «-».
| Достоинства | Недостатки |
| 2-х сторонняя связь позволяет контролировать состояние транспортного средства | Спустя 4-5 лет эксплуатации бюджетные модели грешат случайным срабатыванием |
| Несколько модификаций, которые обеспечивают различный уровень защиты | – |
| Быстрота настройки | – |
| Наличие опций для дальнейшего расширения функционала | – |
| Защита от стороннего вмешательства | – |
| Наличие международного сертификата соответствия | – |
Комментарии производителя свидетельствуют о том, что факты случайного срабатывания обусловлены неправильным обслуживанием или некорректной настройкой. Установку проводят в авторизованном салоне. Самостоятельные попытки выполнить монтаж завершатся повышенным риском угона.
Установку проводят в авторизованном салоне. Самостоятельные попытки выполнить монтаж завершатся повышенным риском угона.
Для установки вам потребуется схема подключения сигнализации Аллигатор, которая должна идти в комплекте к противоугонной системе.
Процесс установки выглядит следующим образом:

Иногда возникает необходимость определить модель противоугонной системы по пульту. Не факт, что вы сможете найти точный ответ, но есть несколько рекомендаций, которые могут помочь вам.
Как узнать модель по брелоку:
За установкой следует пошаговая настройка, потому что Аллигатор сигнализация инструкция от брелка с четырьмя или двумя кнопками, содержит необходимые советы.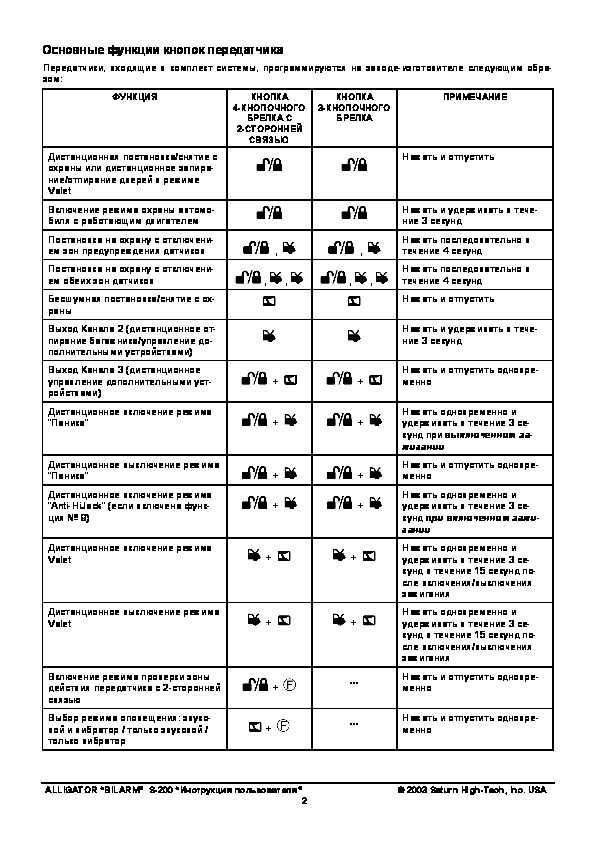 Вставляют ключ в замок зажигания, нажимают и удерживают кнопку Valet. Удержав ее 3 секунды, водитель нажимает ее 3 раза до момента появления звукового сигнала. Финальный шаг заключается в быстром поочередном нажатии на каждую из клавиш, которые расположены на пульте. Дальнейший порядок действий:
Вставляют ключ в замок зажигания, нажимают и удерживают кнопку Valet. Удержав ее 3 секунды, водитель нажимает ее 3 раза до момента появления звукового сигнала. Финальный шаг заключается в быстром поочередном нажатии на каждую из клавиш, которые расположены на пульте. Дальнейший порядок действий:
Выбрав уровень чувствительности, хлопают по капоту.
На заметку! Водитель выбирает значение, при котором машина, с одной стороны, не реагирует на вибрацию, вызванную случайным прохожим, а, с другой стороны, сработает при попытке взлома. После завершения установки фиксируют провода и блок управления так, как это указано в инструкции.
После завершения установки фиксируют провода и блок управления так, как это указано в инструкции.
Водителю останется выбрать один из трех режимов работы. Первый – это «паника», которая запускает звуковое оповещение. Нажимают кнопку открытия/закрытия замка. Повторное нажатие выключит оповещение. Второй режим – это Valet, который отключает автозапуск системы Аллигатор, например, во время ТО или ремонта. Процесс активации выглядит следующим образом:
Третий режим, как было сказано ранее, это Antihijack. Блокировка электрической цепи не позволит вору далеко уехать. Как только система фиксирует проникновение, двигатель оперативно блокируется. Самостоятельное выключение и автозапуск системы Аллигатор, с инструкции как завести с брелка можно узнать детали, позволяет быстро вмешаться в процесс.
Автомобильная сигнализация подходит отечественным и иностранным машинам. Выбирают ее с учетом требуемого уровня защиты, набора опций и характеристик электрической цепи транспортного средства. Эргономичный брелок системы Аллигатор и инструкция к нему позволяет быстро понять функционал. В зависимости от модификации, пульт управления оснащен LED экраном, который отображает параметры работы силового агрегата. Приобретают и устанавливают системы в авторизованном центре. Настройку допустимо выполнить самостоятельно, исходя из фактических условий эксплуатации.
Biol Lett. 2019 июль; 15(7): 20190354.
Опубликовано в сети 3 июля 2019 г. doi: 10.1098/rsbl.2019.0354
, 1 , 3 , 9000 7 4 , 2 , 2 , 5 и 6
Информация об авторе Примечания к статье Информация об авторских правах и лицензиях Отказ от ответственности
Мышцы, обеспечивающие вентиляцию легких, являются ключом к пониманию эволюционных ограничений формы и функций животных. Здесь с помощью электромиографии мы демонстрируем недавно обнаруженную дыхательную функцию подвздошно-реберной мышцы у американского аллигатора ( Alligator mississippiensis ). Подвздошно-реберная мышца активна во время выдоха при дыхании на суше при температуре 28°С, и эта активность опосредована крючковидными отростками позвоночных ребер. Также наблюдалось увеличение мышечной активности во время форсированных выдохов при тревожных вокализациях бедствия. Интересно, что мы не обнаружили никакой дыхательной активности у подвздошно-реберных мышц, когда аллигаторы дышали, погружая свое тело в воду при температуре 18°C, что приводило к снижению частоты дыхания. Подвздошно-реберная мышца — это вспомогательная дыхательная мышца, которую аллигаторы могут использовать для облегчения выдоха при определенных условиях.
Здесь с помощью электромиографии мы демонстрируем недавно обнаруженную дыхательную функцию подвздошно-реберной мышцы у американского аллигатора ( Alligator mississippiensis ). Подвздошно-реберная мышца активна во время выдоха при дыхании на суше при температуре 28°С, и эта активность опосредована крючковидными отростками позвоночных ребер. Также наблюдалось увеличение мышечной активности во время форсированных выдохов при тревожных вокализациях бедствия. Интересно, что мы не обнаружили никакой дыхательной активности у подвздошно-реберных мышц, когда аллигаторы дышали, погружая свое тело в воду при температуре 18°C, что приводило к снижению частоты дыхания. Подвздошно-реберная мышца — это вспомогательная дыхательная мышца, которую аллигаторы могут использовать для облегчения выдоха при определенных условиях.
Ключевые слова: электромиография, дыхание, крючковидные отростки, крокодил
Крокодилы – крупные полуводные рептилии, впервые появившиеся в позднем меловом периоде и имеющие общего предка с птицами. Как и все амниоты, крокодилы используют для вентиляции аспирационный насос [1,2]. У птиц и крокодилов насос создает однонаправленный поток воздуха через легкие [3,4]. Вдох, выдох или и то, и другое являются активными процессами у позвоночных и поэтому могут быть облегчены только за счет мышечной деятельности [5]. Между дыхательными системами птиц и крокодилов много общего; легкие у них многокамерные; внутренние камеры закручиваются по спирали по часовой стрелке и имеют асимметричный характер ветвления [6]. А также есть сходства в топографии внутрилегочного бронха, бронхов [7] и аэродинамических клапанов [4]. Однонаправленный поток воздуха через легкие аллигаторов, подобный наблюдаемому у птиц, скорее всего, присутствовал у базальных архозавров триаса, еще до эволюции птиц [4].
Как и все амниоты, крокодилы используют для вентиляции аспирационный насос [1,2]. У птиц и крокодилов насос создает однонаправленный поток воздуха через легкие [3,4]. Вдох, выдох или и то, и другое являются активными процессами у позвоночных и поэтому могут быть облегчены только за счет мышечной деятельности [5]. Между дыхательными системами птиц и крокодилов много общего; легкие у них многокамерные; внутренние камеры закручиваются по спирали по часовой стрелке и имеют асимметричный характер ветвления [6]. А также есть сходства в топографии внутрилегочного бронха, бронхов [7] и аэродинамических клапанов [4]. Однонаправленный поток воздуха через легкие аллигаторов, подобный наблюдаемому у птиц, скорее всего, присутствовал у базальных архозавров триаса, еще до эволюции птиц [4].
В настоящее время хорошо изучена кинематика грудной клетки при вентиляции у крокодилов [8]. Во время выдоха наружные и парастернально-внутренние межреберья поворачивают ребра внутрь и назад [9], сокращение поперечной мышцы живота перемещает печень вперед, а прямая мышца живота тянет бедра вперед и втягивает живот.
Крючковидные отростки — это выступы, которые отходят назад от проксимальной поверхности позвоночных ребер и являются еще одной общей характеристикой крокодилов и птиц [12]. У птиц эти отростки функционируют как рычаги и участвуют во вдохе и выдохе за счет действия реберно-аппендикулярной и наружной косой мышц соответственно [12]. Морфология крючковидных отростков у птиц варьирует в зависимости от степени специализации к разным формам локомоций [13]. У крокодилов часто упускают из виду наличие крючковидных отростков, поскольку эти отростки хрящевые, что может объяснить их отсутствие в музейных и ископаемых образцах.
Открыть в отдельном окне
( a ) Схема осевой мускулатуры A. mississippiensis , иллюстрирующая подвздошно-реберную мышцу (ic), глубокую наружную косую мышцу живота (eop), прямую мышцу живота (ra) и truncocaudalis (tc). Звездочка указывает на размещение электрода ЭМГ в ic. ( b ) Мышцы были удалены, чтобы выделить крючковидные отростки (стрелки, UCP). Краниальный слева.
mississippiensis , иллюстрирующая подвздошно-реберную мышцу (ic), глубокую наружную косую мышцу живота (eop), прямую мышцу живота (ra) и truncocaudalis (tc). Звездочка указывает на размещение электрода ЭМГ в ic. ( b ) Мышцы были удалены, чтобы выделить крючковидные отростки (стрелки, UCP). Краниальный слева.
Исследовали электромиографическую (ЭМГ) активность подвздошно-реберной мышцы в трех условиях: (1) в покое и дыхании при температуре окружающей среды 28°С, (2) в покое и дыхании с 10-кратно меньшей частотой дыхания при тело погружали в резервуар с водой при температуре 18°С с опорой на передние ноги на блок, удерживая голову в дыхательной камере и (3) во время форсированного выдоха при тревожных вокализациях бедствия при температуре окружающей среды 28°С. Эксперименты проводились на пяти-восьми самках аллигаторов, вылупившихся из яиц, собранных в Луизиане и выращенных в отделении для животных Университета Северного Техаса (средняя масса тела ± s.e. 139).5 ± 150 г; возраст 2 года) содержали в загонах из стекловолокна (0,7 × 2 × 0,7 м) в пресной воде и кормили коммерческим кормом для аллигаторов два раза в неделю.
Анестезию вызывали, помещая голову аллигатора в пластиковую трубку, содержащую ватно-марлевую салфетку, пропитанную изофлураном (Henry Schein Animal Health, Дублин, Огайо, США). Затем трахею интубировали Tygon 9.Трубки 0007®
 После завершения всех экспериментов животных умерщвляли передозировкой пентобарбитона (150 мг/кг -1 , Fatal-Plus ® , Vortech Pharmaceutical, Дирборн, Мичиган, США) и используется для анатомического исследования и подтверждения размещения электродов ЭМГ. Все данные ЭМГ были собраны на оборудовании от Telemetry Research Limited ® (Окленд, Новая Зеландия). С левой стороны у животных путем пальпации кожи над участком локализовали межреберные промежутки между третьим и четвертым грудными ребрами. Затем делают разрез длиной 2–3 см, чтобы обнажить целевую область подвздошно-реберной мышцы. Двойной передатчик биопотенциала (TR40BB) затем пришивали к спине каждого животного примерно в середине позвоночного столба. Затем экранированные ЭМГ-электроды (диаметром 1 мм) туннелировали под кожу от заднего края грудного отдела (2–3 см от места имплантации ЭМГ) до разреза над подвздошно-реберной мышцей. Затем электроды ЭМГ вшивали непосредственно в мышечное брюшко подвздошно-реберной мышцы над крючковидными отростками.
После завершения всех экспериментов животных умерщвляли передозировкой пентобарбитона (150 мг/кг -1 , Fatal-Plus ® , Vortech Pharmaceutical, Дирборн, Мичиган, США) и используется для анатомического исследования и подтверждения размещения электродов ЭМГ. Все данные ЭМГ были собраны на оборудовании от Telemetry Research Limited ® (Окленд, Новая Зеландия). С левой стороны у животных путем пальпации кожи над участком локализовали межреберные промежутки между третьим и четвертым грудными ребрами. Затем делают разрез длиной 2–3 см, чтобы обнажить целевую область подвздошно-реберной мышцы. Двойной передатчик биопотенциала (TR40BB) затем пришивали к спине каждого животного примерно в середине позвоночного столба. Затем экранированные ЭМГ-электроды (диаметром 1 мм) туннелировали под кожу от заднего края грудного отдела (2–3 см от места имплантации ЭМГ) до разреза над подвздошно-реберной мышцей. Затем электроды ЭМГ вшивали непосредственно в мышечное брюшко подвздошно-реберной мышцы над крючковидными отростками.
Дыхание контролировали с помощью датчика давления Validyne ® (модель DP-45-16, Validyne Engineering Corp, Нортридж, Калифорния, США), подключенного к пневмотахометру либо с лицевой маской, изготовленной из модифицированного пластикового стакана объемом 50 мл добавить впускную и выпускную трубки и загерметизировать вокруг головы аллигатора с помощью Bisico ® (ISO4823, тип 3, Bielefelder Dental Silicone GmbH, Германия), когда они находятся в состоянии покоя и дышать, или выпускную трубу дыхательной камеры над резервуаром для воды для дыхание в состоянии покоя при погружении тела в воду (электронный дополнительный материал, рис. S2). В обоих случаях через систему прокачивался воздух со скоростью 500 мл мин −1 . Сигналы бедствия вызывались легким постукиванием по голове аллигатора двумя пальцами и синхронизировались с кривыми ЭМГ путем ручного нажатия на специальный 5-вольтовый триггер, подключенный к PowerLab ®. Необработанные данные ЭМГ подвергались полосовой фильтрации (60–250 Гц), выпрямлению и сглаживанию (треугольное сглаживание Бартлетта) в LabChart ® . В каждом дыхательном цикле обработанный сигнал был разделен на 50 бинов для вдоха и выдоха, с поправкой на продолжительность фазы цикла. В каждом интервале регистрировали среднюю интенсивность ЭМГ-активности (мВ) и интегрированный сигнал, что позволяло количественно оценить начало мышечной активности. Дисперсионный анализ суммы интегрированных ЭМГ по 10 вдохам у каждого животного использовался для проверки того, происходила ли мышечная активность в фазе либо с вдохом, либо с выдохом (электронный дополнительный материал, таблицы S1 и S2).
Сигналы бедствия вызывались легким постукиванием по голове аллигатора двумя пальцами и синхронизировались с кривыми ЭМГ путем ручного нажатия на специальный 5-вольтовый триггер, подключенный к PowerLab ®. Необработанные данные ЭМГ подвергались полосовой фильтрации (60–250 Гц), выпрямлению и сглаживанию (треугольное сглаживание Бартлетта) в LabChart ® . В каждом дыхательном цикле обработанный сигнал был разделен на 50 бинов для вдоха и выдоха, с поправкой на продолжительность фазы цикла. В каждом интервале регистрировали среднюю интенсивность ЭМГ-активности (мВ) и интегрированный сигнал, что позволяло количественно оценить начало мышечной активности. Дисперсионный анализ суммы интегрированных ЭМГ по 10 вдохам у каждого животного использовался для проверки того, происходила ли мышечная активность в фазе либо с вдохом, либо с выдохом (электронный дополнительный материал, таблицы S1 и S2).
Подвздошно-реберная мышца ( a ) представляет собой сегментированную мышцу, которая соединяется с позвонками и проходит поверхностно через позвоночные ребра. Он также занимает межреберные промежутки и прикрепляется к крючковидным отросткам. Крючковатые отростки находятся внутри миосепты, которая сегментирует подвздошно-реберную мышцу, поверхностно и дорсально по отношению к вентральным наружным межреберьям ( b ). Мышечные волокна подвздошно-реберной мышцы берут начало от заднего края позвоночных ребер и охватывают крючковидные отростки, поэтому вполне вероятно, что они будут удалены при любом удалении мускулатуры скелета. Поверхностный ( а ) и глубокая наружная косая мышца проходит через промежуточную и грудинную части ребер, поверхностная часть прикрепляется к фасции, покрывающей подвздошно-реберную мышцу. Слои наружной косой мышцы живота прикрепляются к прямой и каудальной мышцам живота ( a ).
Он также занимает межреберные промежутки и прикрепляется к крючковидным отросткам. Крючковатые отростки находятся внутри миосепты, которая сегментирует подвздошно-реберную мышцу, поверхностно и дорсально по отношению к вентральным наружным межреберьям ( b ). Мышечные волокна подвздошно-реберной мышцы берут начало от заднего края позвоночных ребер и охватывают крючковидные отростки, поэтому вполне вероятно, что они будут удалены при любом удалении мускулатуры скелета. Поверхностный ( а ) и глубокая наружная косая мышца проходит через промежуточную и грудинную части ребер, поверхностная часть прикрепляется к фасции, покрывающей подвздошно-реберную мышцу. Слои наружной косой мышцы живота прикрепляются к прямой и каудальной мышцам живота ( a ).
Мы имплантировали электроды в тело подвздошно-реберной мышцы непосредственно над крючковидными отростками позвоночных ребер ( a , b ). Когда аллигаторы находились в состоянии покоя и дышали со скоростью 13,21 ± 0,77 вдохов в минуту (уд/мин) при температуре окружающей среды 28°C, фазовая активность подвздошно-реберной мышцы коррелировала с выдохом (9). 0030 и ; электронный дополнительный материал, рисунок S1 и таблица S1). Активность подвздошно-реберных мышц не является результатом того, что аллигаторы носят маску (электронный дополнительный материал, рисунок S2). Интересно, однако, что не было фазовой дыхательной активности подвздошно-реберной мышцы, когда аллигаторы находились в покое и дышали, но их тело было погружено в резервуар с водой при температуре 18°C; как при высокой частоте дыхания (4,6 ± 0,92 уд/мин, b ), так и при примерно 10-кратном снижении (1,57 ± 0,46 уд/мин, с ; электронный дополнительный материал, рисунок S3). Подвздошно-реберная мышца также продемонстрировала значительно большую фазовую активность (приблизительно в 20 раз) во время выдохов, связанных с сигнализацией тревоги, по сравнению с дыханием животного в состоянии покоя ( d ; электронный дополнительный материал, рисунок S1 и таблица S2).
0030 и ; электронный дополнительный материал, рисунок S1 и таблица S1). Активность подвздошно-реберных мышц не является результатом того, что аллигаторы носят маску (электронный дополнительный материал, рисунок S2). Интересно, однако, что не было фазовой дыхательной активности подвздошно-реберной мышцы, когда аллигаторы находились в покое и дышали, но их тело было погружено в резервуар с водой при температуре 18°C; как при высокой частоте дыхания (4,6 ± 0,92 уд/мин, b ), так и при примерно 10-кратном снижении (1,57 ± 0,46 уд/мин, с ; электронный дополнительный материал, рисунок S3). Подвздошно-реберная мышца также продемонстрировала значительно большую фазовую активность (приблизительно в 20 раз) во время выдохов, связанных с сигнализацией тревоги, по сравнению с дыханием животного в состоянии покоя ( d ; электронный дополнительный материал, рисунок S1 и таблица S2).
Открыть в отдельном окне
Электромиография (ЭМГ) подвздошно-реберной мышцы. Верхняя кривая на каждой панели представляет собой типичный необработанный необработанный сигнал ЭМГ (мВ), средняя кривая обработана, выпрямленная активность ЭМГ для всех аллигаторов (мВ, полосовой фильтр 60–250 Гц), а нижняя кривая указывает; вдох и выдох для ( a – c ) и обозначает события вокализации сигнала бедствия для ( d ). Все следы принадлежат одному аллигатору. Подвздошно-реберная мышца активна во время выдоха, когда аллигаторы находились в покое и дышали при 28°C ( a ), не проявляла активности ни во время выдоха, ни во время вдоха в состоянии покоя и дышала либо быстро (b ), либо медленнее ( c ). ) при погружении тела в воду с температурой 18°С и демонстрировал заметное усиление активности на выдохе, связанное с тревожными вокализациями ( д ). События вокализации тревоги (нижняя кривая, d ) обозначались ручным нажатием триггера 5 В после каждой вокализации, показанной на нижней кривых, отсюда небольшое смещение, вызванное задержкой времени реакции на событие вокализации тревоги.
Верхняя кривая на каждой панели представляет собой типичный необработанный необработанный сигнал ЭМГ (мВ), средняя кривая обработана, выпрямленная активность ЭМГ для всех аллигаторов (мВ, полосовой фильтр 60–250 Гц), а нижняя кривая указывает; вдох и выдох для ( a – c ) и обозначает события вокализации сигнала бедствия для ( d ). Все следы принадлежат одному аллигатору. Подвздошно-реберная мышца активна во время выдоха, когда аллигаторы находились в покое и дышали при 28°C ( a ), не проявляла активности ни во время выдоха, ни во время вдоха в состоянии покоя и дышала либо быстро (b ), либо медленнее ( c ). ) при погружении тела в воду с температурой 18°С и демонстрировал заметное усиление активности на выдохе, связанное с тревожными вокализациями ( д ). События вокализации тревоги (нижняя кривая, d ) обозначались ручным нажатием триггера 5 В после каждой вокализации, показанной на нижней кривых, отсюда небольшое смещение, вызванное задержкой времени реакции на событие вокализации тревоги.
Понимание эволюции дыхательных структур затруднено, поскольку мягкие ткани, такие как легкие, редко окаменевают. Кроме того, все четвероногие произошли от предков рыб, и расхождение гипаксиальных мышц с локомоторной на дыхательную роль создает возможность для антагонистических функций. Преодоление ограничения Кэрриера, как известен этот конфликт [14,15], привело к эволюции новых вспомогательных дыхательных структур и рекрутированию мышц по всей линии четвероногих.
Наши данные показывают, что подвздошно-реберная мышца, в которую встроены крючковидные отростки, является вспомогательной дыхательной мышцей, способствующей выдоху в периоды повышенной частоты дыхания за счет перемещения грудной клетки внутрь и назад. Хорошо известно, что, как было обнаружено здесь, частота дыхания у крокодилов напрямую коррелирует с температурой тела и увеличением частоты дыхания [16]. Кроме того, известно, что у крокодилов изменения в интенсивности мышечной деятельности и задействовании различных мышц связаны с различиями в частоте дыхания. Например, интенсивность упражнений влияет на механику дыхания [17]. Когда крокодилы отдыхают или ходят, диафрагмальный вклад вносит ограниченный вклад. Однако по мере увеличения уровня и интенсивности усилий (при плавании и скоростной ходьбе) основной вклад вносят диафрагмальные мышцы [17]. Подвздошно-реберная мышца также функционирует при постуральной поддержке и сгибании туловища во время передвижения у некоторых рептилий [18]. Описанная здесь роль вспомогательной дыхательной мышцы не исключает двойной роли подвздошно-реберной мышцы в поддержке или сагиттальном изгибе тела во время передвижения у аллигатора; однако это еще предстоит определить. У птиц, например, реберно-аппендикулярная мышца играет двойную роль во время дыхания и передвижения [12]. Будущие эксперименты по изучению того, были ли какие-либо изменения в мышечной активности вдоль рострокаудальной оси этой мышцы у крокодилов, также могут пролить новый свет на эту потенциальную двойную роль. Важность задействования подвздошно-реберной мышцы для облегчения выдоха была подтверждена относительным увеличением мышечной активности во время вокализации.
Например, интенсивность упражнений влияет на механику дыхания [17]. Когда крокодилы отдыхают или ходят, диафрагмальный вклад вносит ограниченный вклад. Однако по мере увеличения уровня и интенсивности усилий (при плавании и скоростной ходьбе) основной вклад вносят диафрагмальные мышцы [17]. Подвздошно-реберная мышца также функционирует при постуральной поддержке и сгибании туловища во время передвижения у некоторых рептилий [18]. Описанная здесь роль вспомогательной дыхательной мышцы не исключает двойной роли подвздошно-реберной мышцы в поддержке или сагиттальном изгибе тела во время передвижения у аллигатора; однако это еще предстоит определить. У птиц, например, реберно-аппендикулярная мышца играет двойную роль во время дыхания и передвижения [12]. Будущие эксперименты по изучению того, были ли какие-либо изменения в мышечной активности вдоль рострокаудальной оси этой мышцы у крокодилов, также могут пролить новый свет на эту потенциальную двойную роль. Важность задействования подвздошно-реберной мышцы для облегчения выдоха была подтверждена относительным увеличением мышечной активности во время вокализации. Аллигаторы имеют большой вокальный репертуар, который зависит от активного контроля движений гортани и выдоха [18]. Использование подвздошно-реберной мышцы для облегчения дыхания характерно не только для аллигаторов; у подвязочной змеи соскальзывание подвздошно-реберной мышцы, двуглавой мышцы ретрактора кости, способствует вдоху [19].]. Однако более распространенным является вовлечение гипаксиальных мышц для облегчения дыхания у других рептилий [15,20] или млекопитающих [21]. Отсутствие экспираторной активности подвздошно-реберной мышцы, когда аллигаторы находились в состоянии покоя и дышали либо относительно медленно, либо быстро при погружении тела, вероятно, связано со снижением скорости метаболизма при более низкой температуре тела в воде. В условиях сниженной аэробной потребности гидростатическое давление, действующее на брюшную полость, которое смещает печень дальше вглубь грудной клетки, может способствовать пассивному выдоху [9].].
Аллигаторы имеют большой вокальный репертуар, который зависит от активного контроля движений гортани и выдоха [18]. Использование подвздошно-реберной мышцы для облегчения дыхания характерно не только для аллигаторов; у подвязочной змеи соскальзывание подвздошно-реберной мышцы, двуглавой мышцы ретрактора кости, способствует вдоху [19].]. Однако более распространенным является вовлечение гипаксиальных мышц для облегчения дыхания у других рептилий [15,20] или млекопитающих [21]. Отсутствие экспираторной активности подвздошно-реберной мышцы, когда аллигаторы находились в состоянии покоя и дышали либо относительно медленно, либо быстро при погружении тела, вероятно, связано со снижением скорости метаболизма при более низкой температуре тела в воде. В условиях сниженной аэробной потребности гидростатическое давление, действующее на брюшную полость, которое смещает печень дальше вглубь грудной клетки, может способствовать пассивному выдоху [9].].
Крючковидные отростки могут быть ключевыми вспомогательными дыхательными структурами в линии четвероногих. Они также встречаются у динозавров-манирапторов, включая велоцирапторов и овирапторов, и могут играть роль в механике дыхания, подобной птичьей, у этих животных [22]. Некоторые четвероногие динозавры также обладали аналогичными межреберными пластинами, которые, как считается, играют роль в дыхании или передвижении [23]. Тенденции двуногости и повышенной беглости, наблюдаемые у динозавров [24], также развились в линии крокодилов [25], указывая на то, что у самых ранних архозавров существовало необходимое разделение вентиляции и передвижения. За пределами линии архозавров у ранних амфибий также сообщалось о широких пластинчатых крючковидных отростках 9.0030 Eyrops и Dissorophus [26]. Интересно, что, учитывая наши находки для аллигатора, подвздошно-реберная мышца простиралась на ребра у Eryops [27]. Крючковатые отростки отмечены также у одного вида вымершей лягушки Prosalirus biti [28]. Любая потенциальная роль крючковидных желез в дыхании амфибий неизвестна, хотя выдох является пассивным у современных бесхвостых и пассивным или активным у уроделей [2].
Они также встречаются у динозавров-манирапторов, включая велоцирапторов и овирапторов, и могут играть роль в механике дыхания, подобной птичьей, у этих животных [22]. Некоторые четвероногие динозавры также обладали аналогичными межреберными пластинами, которые, как считается, играют роль в дыхании или передвижении [23]. Тенденции двуногости и повышенной беглости, наблюдаемые у динозавров [24], также развились в линии крокодилов [25], указывая на то, что у самых ранних архозавров существовало необходимое разделение вентиляции и передвижения. За пределами линии архозавров у ранних амфибий также сообщалось о широких пластинчатых крючковидных отростках 9.0030 Eyrops и Dissorophus [26]. Интересно, что, учитывая наши находки для аллигатора, подвздошно-реберная мышца простиралась на ребра у Eryops [27]. Крючковатые отростки отмечены также у одного вида вымершей лягушки Prosalirus biti [28]. Любая потенциальная роль крючковидных желез в дыхании амфибий неизвестна, хотя выдох является пассивным у современных бесхвостых и пассивным или активным у уроделей [2]. Птерозавры имеют аналогичные структуры, sternocostapophyses, которые, как считается, имеют сходную функцию с крючковидными отростками во время движений ребер и уменьшают работу дыхания [29].].
Птерозавры имеют аналогичные структуры, sternocostapophyses, которые, как считается, имеют сходную функцию с крючковидными отростками во время движений ребер и уменьшают работу дыхания [29].].
Демонстрация дополнительной дыхательной функции подвздошно-реберной мышцы, опосредованной крючковидными отростками, предполагает, что гибкость механики дыхания у крокодилов, связанная с изменениями потребности в дыхании, вероятно, развилась у базальных архозавров.
Дополнительные рисунки:
Нажмите здесь для просмотра. (760K, docx)
Мы благодарим Дерека Нельсена, Джастина Коннера и Жанну Кроссли за помощь в разведении и дрессировке аллигаторов.
Дж.Р.К. задумал идею и разработал исследование. J.R.C., K.A.R.R., P.G.T., W.I.S., R.J.B., R.M.E. и D.A.C.II собрали, интерпретировали и проанализировали данные. Р.М.Э. облегченный сбор яиц аллигатора. Все авторы подготовили и отредактировали рукопись. Все авторы соглашаются нести ответственность за содержание и одобряют окончательную версию этой рукописи.
Авторы заявляют, что у них нет конкурирующих интересов.
Исследование финансировалось BBSRC (BB/I021116/1) и NSF (IBN IOS-0845741).
1. Бернер Р.А., ВанденБрукс Дж.М., Уорд П.Д.. 2007. Кислород и эволюция. Наука 316, 557–558. ( 10.1126/science.1140273) [PubMed] [CrossRef] [Google Scholar]
2. Брейнерд Э.Л. 2009. Новые взгляды на эволюцию механизмов вентиляции легких у позвоночных. Эксп. биол. В сети 4, 11–28. (doi:10/1007/s00898-999-0002-1) [Google Scholar]
3. Scheid P, Piiper J. 1971. Прямое измерение пути выдыхаемого газа в легких уток. Дыхание Физиол. 11, 308–314. ( 10.1016/0034-5687(71)
-1) [PubMed] [CrossRef] [Google Scholar]
4. Farmer CG, Sanders K. 2010. Однонаправленный поток воздуха в легких аллигаторов. Наука 327, 338–340. ( 10.1126/science.1180219) [PubMed] [CrossRef] [Google Scholar]
5. Klein W, Codd JR.
2010.
Дыхание и передвижение: сравнительная анатомия, морфология и функции. Дыхание Физиол. Нейробиол.
173, С26–С32. (10.1016/j.resp.2010.04.019) [PubMed] [CrossRef] [Google Scholar]
(10.1016/j.resp.2010.04.019) [PubMed] [CrossRef] [Google Scholar]
6. Perry SF. 1990. Стратегия газообмена у нильского крокодила: морфометрическое исследование. Дж. Комп. Физиол. Б 159, 761–769. ( 10.1007/BF00691722) [CrossRef] [Google Scholar]
7. Перри С.Ф. 1998. Легочная система: легкие: сравнительная анатомия, функциональная морфология и эволюция. В биологии рептилий, том. 19 (ред. Ганс С., Гонт А.С.), стр. 1–92. Итака, Нью-Йорк: Academic Press. [Google Scholar]
8. Броклхерст Р.Дж., Мориц С., Кодд Дж.Р., Селлерс В.И., Брейнерд Э.Л. 2017. Кинематика ребер во время вентиляции легких у американского аллигатора ( Alligator mississippiensis ): анализ XROMM. Дж. Эксп. биол. 220, 3181–3190. ( 10.1242/jeb.156166) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Gans C, Clark B. 1976 год. Исследования вентиляции Caiman crocodilus (Crocodilia: Reptilia). Дыхание Физиол. 26, 285–301. ( 10.1016/0034-5687(76)-3) [PubMed] [CrossRef] [Google Scholar]
10. Farmer CG, Carrier DR.
2000.
Тазовая аспирация у американского аллигатора ( Alligator mississippiensis ). Дж. Эксп. биол.
203, 1679–1687. [PubMed] [Академия Google]
Farmer CG, Carrier DR.
2000.
Тазовая аспирация у американского аллигатора ( Alligator mississippiensis ). Дж. Эксп. биол.
203, 1679–1687. [PubMed] [Академия Google]
11. Перри С.Ф. 1988 год. Функциональная морфология легких нильского крокодила Crocodylus niloticus : нереспираторные параметры. Дж. Эксп. биол. 134, 99–117. [Google Scholar]
12. Кодд Дж. Р., Боггс Д. Ф., Перри С. Ф., Кэрриер Д. Р. 2005. Активность трех мышц, связанных с крючковидными отростками гигантского канадского гуся Branta canadensis maximus . Дж. Эксп. биол. 208, 849–857. ( 10.1242/jeb.01489) [PubMed] [CrossRef] [Google Scholar]
13. Tickle PG, Ennos AR, Lennox LE, Perry SF, Codd JR. 2007. Функциональное значение крючковидных отростков у птиц. Дж. Эксп. биол. 210, 3955–3961. ( 10.1242/jeb.008953) [PubMed] [CrossRef] [Google Scholar]
14. Carrier DR.
1991.
Конфликт в гипоаксиальной мышечно-скелетной системе: документирование эволюционного ограничения. интегр. Комп. биол.
31, 644–654. ( 10.1093/icb/31.4.644) [CrossRef] [Google Scholar]
Комп. биол.
31, 644–654. ( 10.1093/icb/31.4.644) [CrossRef] [Google Scholar]
15. Перевозчик DR. 1989. Вентиляционное действие гипаксиальных мышц ящерицы Iguana iguana : функция медленных мышц. Дж. Эксп. биол. 143, 435–457. [PubMed] [Google Scholar]
16. Маннс С.Л., Эванс Б.К., Фраппелл П.Б. 1998. Влияние температуры окружающей среды, гипоксии и гиперкапнии на характер дыхания морских крокодилов ( Crocodylus porosus ). Дж. Эксп. биол. 71, 267–273. ( 10.1086/515913) [PubMed] [CrossRef] [Google Scholar]
17. Munns SL, Owerkowicz T, Andrewartha SJ, Frappell PB. 2012. Вспомогательная роль диафрагмальной мышцы в вентиляции легких у эстуарного крокодила Crocodylus porosus . Дж. Эксп. биол. 215, 845–852. ( 10.1242/jeb.061952) [PubMed] [CrossRef] [Google Scholar]
18. Риде Т., Ли З., Токуда И.Т., Фармер К.Г.
2015.
Функциональная морфология гортани миссисипского аллигатора с последствиями для вокала. Дж. Эксп. биол.
218, 991–998. ( 10.1242/jeb.117101) [PubMed] [CrossRef] [Google Scholar]
( 10.1242/jeb.117101) [PubMed] [CrossRef] [Google Scholar]
19. Розенберг HI. 1973. Функциональная анатомия легочной вентиляции у подвязочной змеи, Thamnophis elegans . Дж. Морфол. 140, 171–184. ( 10.1002/jmor.1051400205) [PubMed] [CrossRef] [Google Scholar]
20. Baumel JJ, Wilson JA, Bergren DR. 1990. Вентиляционные движения таза и хвоста птиц: функции мышц хвостового отдела голубя. Дж. Эксп. биол. 151, 263–277. [PubMed] [Google Scholar]
21. Дебан С.М., Carrier DR. 2002. Активность гипоаксиальных мышц при беге и дыхании у собак. Дж. Эксп. биол. 205, 1953–1967. [PubMed] [Google Scholar]
22. Codd JR, Manning PL, Perry SF, Norell MA. 2008. Птичье дыхание у динозавров-манирапторов. проц. Р. Соц. Б 275, 157–161. ( 10.1098/rspb.2007.1233) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
23. Boyd CA, Cleland TP, Navas F.
2011.
Остеогенез, гомология и функция межреберных пластин у птицетазовых динозавров (Tetrapoda, Sauropsida). Зооморфология
130, 303–313. ( 10.1007/s00435-011-0136-x) [CrossRef] [Google Scholar]
Зооморфология
130, 303–313. ( 10.1007/s00435-011-0136-x) [CrossRef] [Google Scholar]
24. Persons WS, Currie PJ. 2017. Функциональное происхождение двуногости динозавров: совокупные данные о склонных к двуногим рептилиях и млекопитающих, не склонных к наклону. Дж. Теор. биол. 420, 1–7. ( 10.1016/j.jtbi.2017.02.032) [PubMed] [CrossRef] [Google Scholar]
25. Готье Дж., Несбитт С.Дж., Шахнер Э.Р., Бевер Г.С., Джойс В.Г. 2011. Двуногий стволовой крокодил Poposaurus gracilis : определение функции окаменелостей и инноваций в передвижении архозавров. Бык. Пибоди Мус. Нац. История 52, 107–126. ( 10.3374/014.052.0102) [CrossRef] [Google Scholar]
26. Clack JA. 2002. Набирает силу эволюция земности. В книге «Завоевание земли: происхождение и эволюция четвероногих», стр. 278–332. Блумингтон, Индиана: Издательство Индианского университета. [Академия Google]
27. Олсон Э.К.
1936 год.
Спинная осевая мускулатура некоторых примитивных пермских четвероногих. Дж. Морфол.
59, 265–311. ( 10.1002/jmor.10505) [CrossRef] [Google Scholar]
Морфол.
59, 265–311. ( 10.1002/jmor.10505) [CrossRef] [Google Scholar]
28. Шубин Н.Х., Дженкинс Дж.Р. 1995. Раннеюрская прыгающая лягушка. Природа 377, 49–52. ( 10.1038/377049a0) [CrossRef] [Google Scholar]
29. Claessens LPAM, O’Connor PM, Unwin DM. 2009. Эволюция дыхания способствовала возникновению полета птерозавров и воздушного гигантизма. ПЛОС ОДИН 4, 1–8. ( 10.1371/journal.pone.0004497) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
30. Риттер Д. 1995. Эпаксиальная мышечная функция во время передвижения у ящерицы ( Varanus salvator ) и предложение ключевого нововведения в осевой опорно-двигательной системе позвоночных. Дж. Эксп. биол. 198, 2477–2490. [PubMed] [Google Scholar]
Джимми Бентли , Patch Staff
Ниже приведены выдержки из журнала полиции Стаутона. Аресты не указывают на обвинительные приговоры:
27 июня
8:28 утра. Полиция Стаутона получила звонок из Центра судьи Роттенберга по поводу студента, выпрыгнувшего из окна. Партия находилась.
17:38 — Звонивший сообщил, что нашел потерянный ноутбук. Он был передан в полицейский участок и возвращен владельцу.
28 июня
9:40 — Поступило сообщение о магазинной краже в магазине Stop and Shop. По словам звонившего, кто-то пытался украсть морепродукты на сумму 170 долларов. Инцидент находится на стадии дальнейшего расследования.
16:10 — В полицию поступил сигнал о фейерверке, предположительно выпущенном из машины на улице Кантон. Рассматриваемый автомобиль имеет номерной знак штата Массачусетс 452TN6.
Рассматриваемый автомобиль имеет номерной знак штата Массачусетс 452TN6.
29 июня
1:38 утра — Полиция Стаутона арестовала 21-летнего Брендона Амадо по обвинению в хранении наркотиков. Автомобиль Амадо был остановлен после того, как он не подал сигнал на повороте на Бродвее.
23:09 — В полицию поступил звонок о том, что расставание мужчины и женщины вызвало беспорядки в ресторане Smokey Bones Barbecue.
По словам звонившего, группа спотыкалась на ходу и казалась пьяной, когда возвращалась к своей машине. Сообщается, что группа уехала на более новой модели Mercedes с номерным знаком 5LC.
30 июня
2:06 утра — Полиция Шарона позвонила в Стаутон с просьбой помочь с остановкой автомобиля.
9:40 — Группа пришла в отделение полиции и сообщила о краже личных данных.
20:03 — Служба контроля за животными расследовала сообщение об аллигаторе возле Taj Estates. Расследование подтвердило, что предполагаемый аллигатор был просто приманкой.